
Рефераты по международному публичному праву
Рефераты по международному частному праву
Рефераты по международным отношениям
Рефераты по культуре и искусству
Рефераты по менеджменту
Рефераты по металлургии
Рефераты по муниципальному праву
Рефераты по налогообложению
Рефераты по оккультизму и уфологии
Рефераты по педагогике
Рефераты по политологии
Рефераты по праву
Биографии
Рефераты по предпринимательству
Рефераты по психологии
Рефераты по радиоэлектронике
Рефераты по риторике
Рефераты по социологии
Рефераты по статистике
Рефераты по страхованию
Рефераты по строительству
Рефераты по таможенной системе
Сочинения по литературе и русскому языку
Рефераты по теории государства и права
Рефераты по теории организации
Рефераты по теплотехнике
Рефераты по технологии
Рефераты по товароведению
Рефераты по транспорту
Рефераты по трудовому праву
Рефераты по туризму
Рефераты по уголовному праву и процессу
Рефераты по управлению
Курсовая работа: Технохимическая характеристика тихоокеанской трески
Курсовая работа: Технохимическая характеристика тихоокеанской трески
Технохимическая характеристика тихоокеанской трески
Введение
Трескоподобные имеют стройное тело, с тремя спинными и двумя анальными плавниками. Хвостовой плавник у них хорошо обособлен, усечен или с выемкой по заднему краю. Икринки не содержат жировой капли, брюшные плавники у личинок не удлинены. Наряду с придонными видами имеются среди них и виды, держащиеся в толще воды и у поверхности [2].
Треска (род Gadus) – очень многочисленный род семейства, с широким ареалом, охватывающим бореальную (умеренную) область Атлантического океана, образует несколько видов и значительное число рас.
Имеет три спинных и два анальных плавника, крупную голову с большим конечным ртом. Усик хорошо развит. Окраска сильно варьирует: цвет спины зеленый или желтовато-коричневый, иногда бурый, с многочисленными мелкими желтовато-коричневыми пятнами. Бока того же тона, более светлые, брюхо желтоватое или белое. Боковая лилия светлая, хорошо заметна на протяжении всего тела, образует небольшой изгиб над грудным плавником.
В Атлантическом океане треска распространена от мыса Код и Бискайского залива до Гренландии, Шпицбергена и Новой Земли (рис. 150). Треска – эврифаг, использующий все виды пищи от планктона до сравнительно крупных рыб.
Очень существенно отличается от атлантической трески и ее подвидов, живущих в морях Атлантического океана, тихоокеанская треска. Она представляет собой особый вид (Gadus macrocephalus), нередко рассматриваемый как подвид (Gadus morhua macrocephalus).
Тихоокеанская треска, в противоположность атлантической и всем подвидам и расам трески атлантического бассейна, имеет не плавучую пелагическую икру, а донную, прилипающую. Она распространена от Берингова пролива до Желтого моря по азиатской стороне и до Орегона вдоль американских берегов.
Наваги (Eleginus) – прибрежные холодолюбивые рыбы северных и восточных морей, распространенные на восток от Мурмана и в северной части Тихого океана. Северная навага (Eleginus navaga) распространена в Белом, Печорском (юго-восточная часть Баренцева), Карском морях, от Восточного Мурмана до Обской губы. Дальневосточная навага, или вахня (Eleginus gracilis), распространена в морях северной части Тихого океана от Берингова пролива до Кореи на западе и до острова Ситки (Аляскинский залив) у американского побережья. Есть она и в Чукотском море по обе стороны от Берингова пролива. Обычна у берегов Берингова, Охотского и Японского морей. Входит в устья рек и в озера.
Путассу (род Micromesistius, два вида) – обитатель пелагиали, питающийся планктоном. Путассу распространена в северной и юго-западной частях Атлантического океана и на юго-западе Тихого океана, в тропической области ее нет. Северная путассу (Micromesistius poutassou) может достигать в длину 47см, но обычная длина ее 30–35см. Она распространена преимущественно в северо-восточных водах Атлантического океана от западной части Средиземного моря до Исландии, Шпицбергена и западных частей Баренцева моря. Есть она и в северо-западных водах от Новой Шотландии и южного склона Большой Ньюфаундлендской банки до Южной Гренландии. Южная путассу (Micromesistius australis) несколько крупнее северной, достигает в длину 50– 55см и массы 1–1,2 кг, отличается лучшей упитанностью. Она распространена в водах склона Патагонского шельфа от Ла-Платы до Южных Оркнейских островов, а также в подах Новозеландского плато, к юго-востоку от Новой Зеландии [4].
Минтай (Theragra chalcogramma) распространен в морях северной части Тихого океана от Берингова пролива до Кореи и Северной Калифорнии, обычно встречаясь при низкой температуре воды, преимущественно от 2 до 9°С.
Пикша (Melanogrammus aeglefinus) распространена только в северных водах Атлантического океана, у берегов Европы, вокруг Исландии и у Северной Америки. По величине улова пикша занимает третье место среди тресковых рыб, следуя за треской и минтаем и давая улов до 1/2 –3/4 млн. т в год [2].
1. Краткие сведения по биологии тихоокеанской трески
Тихоокеанская треска, Восточная треска - Yadus macrocephalus Tilesius; англ. Pacific cod; яп. Madara; кор. Dae-gu.

Тихоокеанская треска имеет вальковатое тело, широкую голову и длинный усик на нижней челюсти. Голова у нее относительно крупнее и шире, чем у атлантической трески. Она сильно отличается от атлантической трески строением рожкообразных выростов переднего конца плавательного пузыря, которые у нее гораздо короче, чем у атлантической. Окраска спины и боков зеленовато-серая, брюхо светлое. Спинных плавников 3, анальных-2. Боковая линия сплошная, светлая, тянется до конца основания третьего спинного плавника.
В водах Приморья треска встречается повсеместно. Широко представлена в Японском, Охотском и Беринговом морях, а также у берегов Северной Америки на юг до Орегона, в тихоокеанских водах Японии, в Желтом море, у берегов Кореи и Китая.
Тихоокеанская треска достигает довольно крупных размеров - длины 125см и массы 24кг. Живет до 15 лет. В промысловых уловах преобладают особи в возрасте 4-8 лет, длиной 45-70см и массой 2,0-4,5кг. Придонный стайный вид. Обитает в придонных горизонтах на шельфе и у верхней кромки материкового склона в водах с температурой от -1,5 до +18 °С. Максимальные концентрации отмечаются в водах с температурой от 0 до 5°С. Впервые тихоокеанская треска созревает в возрасте 4-6 лет при длине 40-50 см. Нерест в водах зал. Петра Великого проходит в феврале-апреле над глубинами 100-150 м, в Северном Приморье - над большими глубинами. Плодовитость составляет 7,0-9,0 млн. икринок. Икра донная, мелкая, диаметром 1,25-1,30 мм, развивается в придонных слоях. Инкубационный период длиться от 10 до 20 дней в зависимости от температуры воды. После нереста треска мигрирует к прибрежному мелководью с глубинами 20-70 м, где интенсивно питается. Будучи холодолюбивой рыбой, тихоокеанская треска после прогрева прибрежных вод отходит от берегов на глубины 80-120 м и более, где держится весь летний период. В осенне-зимний сезон значительных перемещений она не совершает, обитая в зоне глубин 140-250 м. Зимой может иногда выходить и на меньшие глубины, встречаясь изредка. в уловах вентерей, выставляемых в Амурском заливе на навагу. Треска Северного Приморья имеет сходный сезонный тип миграции. Средняя длина ее в промысловых скоплениях составляет 65-75 см.
Тихоокеанская треска характеризуется широким спектром потребляемых пищевых объектов, в число которых входят различные группы беспозвоночных животных и рыб. Неполовозрелые особи (длиной менее 40 см) питаются преимущественно мелкими донными и придонными ракообразными, червями, молодью бычков, камбал и других рыб. Крупная треска питается в основном рыбой - минтаем, сельдью, песчанкой, камбалой, а также кальмарами и осьминогами.
Тихоокеанская треска - важная промысловая рыба. Ее основные скопления приурочены к шельфовым водам Северного Приморья. Лов трески возможен круглый год, однако в Северном Приморье, между м. Олимпиады и б. Нельма, для удобного и ярусного промысла наиболее результативны июль-август, когда треска откармливается вблизи берегов на глубинах 30-75 м. Запасы тихоокеанской трески в водах Приморья в настоящее время недоиспользуются. В 1985-1990 гг. вылов колебался в пределах 1,2-2,5 тыс. т, в 90-е гг. - в пределах 0,5-1,2 тыс.т [1].
2. Краткие сведения о способах лова, транспортирования и хранения трески до обработки
Важная промысловая рыба. Ее основные скопления приурочены к шельсровым водам Северного Приморья, Добывается крючковыми снастями (ярусами и удочками) и тралящими орудиями лова (снюрреводами и тралами). При подходах к берегу попадается в ставные невода. Следует учесть, что при использовании крючковых орудии лова добывается треска более крупных размеров и им нужно отдавать предпочтение перед тралами. Лов трески возможен круглый год, однако в Северном Приморье между м. Олимпиады и б. Нельма, для удобного и ярусного промысла наиболее результативны июль-август, когда треска откармливается вблизи берегов на глубинах 30-75 м. Запасы трески в водах Приморья недоиспользуются. В последнее десятилетие её вылов здесь колебался в пределах 0,35-2,20 тыс. т в год.
Трал представляет собой орудие лова в виде мешка, который буксируют в толще воды или у дна. Тралами ловят косячную и относительно разреженную рыбу в море до глубин 2000— 2500 м и во внутренних водоемах (в основном в озерах и водохранилищах).
Объектами тралового лова являются треска, пикша, морской окунь, камбала, палтус, хек, сардина, скумбрия, ставрида, сельдь, мерлуза, морской карась и т. д. Кроме того, тралами ловят нерыбные объекты — креветок, криля, кальмаров.
Траловый лов широко применяют во всех странах с развитой рыбной промышленностью — в России, Японии, США, Норвегии, Германии и т.д. Он дает примерно 2/3 мирового улова рыбы и около 3/4 добычи рыбы в России.
Широкому распространению тралового лова способствуют его универсальность, высокая активность, автономность, сравнительная простота механизации и автоматизации, возможность полной или частичной переработки рыбы на судах, высокая производительность и экономическая эффективность.
Орудия и способы тралового лова отличаются значительным разнообразием.
По способу горизонтального раскрытия тралы делят на распорные, бимтралы и близнецовые. У распорных тралов горизонтальное раскрытие обеспечивает распорная сила траловых досок, у бимтралов — специальный брус-бим: Близнецовыми тралами работают с двух судов (без траловых досок), и их раскрытие обеспечивают поперечные составляющие натяжения ваеров.
В зависимости от горизонта хода тралы делят на донные, придонные, разноглубинные и универсальные. Донные тралы предназначены для работы по дну водоема, придонные—в непосредственной близости от него, разноглубинные— в толще воды. Универсальные тралы способны работать в донном, придонном и разноглубинном вариантах.
По конструкции сетной части тралы делятся на двухпластные, четырехпластные и многопластные в зависимости от числа пластин, из которых сшит трал.
По материалу оболочки они разделяются на сетные и канатные тралы. В сетных тралах вся оболочка изготовлена из сетного полотна, а в канатных — передняя часть оболочки трала образована канатными элементами. Известны также сетные тралы с канатными крыльями.
По количеству буксировочных тросов при тралении различают тралы одно-, двух- и четырехваерные.
Различают тралы для работы с борта и с кормы, с применением и без применения физических средств интенсификации лова, тралы с гидромеханизацией и без гидромеханизации.
Обычно судно работает одновременно одним тралом. Однако известны способы тралового лова, когда с одного судна одновременно ловят несколькими тралами или сдвоенными тралами.
Траловая система и ее характеристики
Система - это совокупность взаимосвязанных объектов реального мира, выполняющих общую функцию и являющихся предметом исследования. Например, траловая система (ТС) - это совокупность таких объектов, как промысловое судно 1, ваерные лебедки 2, ваерные блоки 3, ваеры 4, траловые доски 5, кабели 6, сетная оболочка трала 7, оснастка гидродинамическим щитком (ГДЩ) 8 верхней подборы, распределенный груз (якорные цепи) 9 и сосредоточенный груз (грузы-углубители) 10 нижней подборы, выполняющих общую функцию лова рыбы тралом (рис. 1).
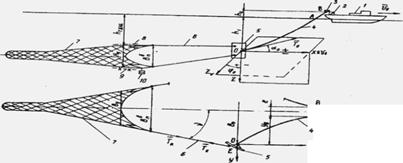
Рисунок 1. Траловая рыболовная система
Траловая система предназначена для лова гидробионтов (рыб, кальмаров, креветок, криля) тралами.
Трал - отцеживающее орудие промышленного рыболовства, представляющее собой сетную оболочку с системами остропки и оснастки, которые обеспечивают его прочность, горизонтальное и вертикальное раскрытия. Трал состоит из двух основных частей: передней - для концентрации и направления рыбы, и задней (мешка) - для отбора рыб промысловых размеров (селекции), их удержания и накопления .
Различают донные и разноглубинные тралы. Донные используют для лова донных (камбалы, палтуса, ската) и придонных (трески, хека, угольной рыбы, морского окуня) рыб, ракообразных (креветок) и других, разноглубинные - для лова пелагических рыб и ракообразных (сельди, сардины, скумбрии, ставриды, минтая, тунца, мавроликуса, криля).
В связи со значительной порчей донными тралами кормовой базы рыб (бентоса) в настоящее время они вытесняются универсальными тралами, позволяющими ловить донных, придонных и пелагических рыб и беспозвоночных.
Эффективность лова рыбы тралами в основном определяется; скоростью траления У8, вертикальным Н;. и горизонтальным Ву раскрытиями устья трала, углом атаки сетной оболочки а n.
Вертикальное раскрытие трала обеспечивается оснасткой его верхней и нижней подбор. Верхняя подборка оборудована шаровыми поплавками (кухтылями) или гидродинамическими щитками (ГДЩ) из жестких (пластмассы, дюралюминия) или гибких (брезента, прорезиненной ткани) материалов; нижняя - якорными цепями для разноглубинного трала или грунтропом для донного трала. Иногда нижнюю подбору донных тралов оснащают щитками из гибких материалов.
Основные части трала и их назначение
Трал состоит из сетной оболочки, ее остропки, оснастки подбор и кабелей. Сетная оболочка трала служит для отцеживания рыб из водной среды. Она состоит из крыльев, мотни, мешка (рис. 10). Крылья позволяют равномерно через подборы передавать нагрузку от мотни к кабелям, также выполняют залавливающие функции. Мотня служит для буксировки мешка, отпугивания рыб и направления их в мешок. При помощи мешка осуществляется отбор рыб промысловых размеров - селекция, их удержание и накопление.
Остропка сетной оболочки включает систему канатов (верхнюю, нижнюю и боковые подборы, сборочные топенанты, пожилины), обеспечивающую общую прочность сетной оболочки. Остропка передает усилия от сетной оболочки на кабели.
Кабели соединяют трал с траловыми досками. Их использование позволяет увеличить зону облова, так как траловые доски и кабели, отпугивая рыб, заставляют их перемещаться к оси трала, концентрироваться в его устье.
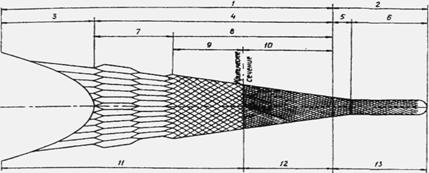
Рис. 10. Сетная оболочка трала (СО):
1 - передняя часть СО; 2 - задняя часть СО (мешок); 3 - крылья: 4 - мотня; 5-коническая часть мешка; 6 - цилиндрическая часть мешка; 7 - канатная часть мотни; 8 - делевая часть мотни; 9 - крупноячейная часть мотни; 10 - мелкоячейная часть мотни; 11 - отпугивающая часть СО; 12 - направляющая часть СО; 13 - аккумулирующая улов часть СО
Залавливающая зона - это часть пространства, ограниченная сетной оболочкой. Она делится на три подзоны: отпугивающую, направляющую и накапливающую улов. Главной функцией отпугивающей части является отпугивание гидробионтов от сетной оболочки трала. Эта подзона охватывает крылья, канатную и крупноячейную части мотни (а > 200 мм). Направляющая подзона - это область, ограниченная мелкоячейной частью мотни. Ее назначение - направить рыбу в мешок трала и не дать ей возможности выйти через ячею оболочки. Границей этих двух подзон является критическое сечение трала. За ним характер поведения рыбы качественно меняется, так как она воспринимает трал как опасность и пытается выйти из него. Это объясняется достижением в этом сечении критической концентрации рыб рсф и подходом их к сетной оболочке ближе, чем на дистанцию реагирования на сетное полотно Вгн.
Положение критического сечения зависит от скорости траления, плотности и размеров облавливаемых стай. При проектировании тралов критическое сечение фиксируется тем, что перед ним сетная оболочка трала выполняется из канатов и крупноячейной дели, за ним - из мелкоячейной дели.
Зимой треска может иногда выходить на меньшие глубины, встречаясь в уловах вентерей, выставляемых в Амурском заливе на навагу. Вентери относятся к стационарным орудиям рыболовства - ловушкам закрытого типа. Вентери состоят из трех конструктивных элементов: направляющего устройства, ловушки, кутка. Направляющее устройство состоит из сетного крыла и открылков (рис. 2). Ловушка представляет собой объемную сетную конструкцию, внутри которой ставятся входные устройства - усынки в виде усеченного конуса. Как правило, количество усынков в вентере составляет 2-3. По конструкции ловушки вентери подразделяются на два типа: кательные и бескательные. Кательные представляют собой сетной цилиндр-бочку, который монтируется на кателях - деревянных обручах. У бескательных вентерей каркас ловушки делается из канатов и имеет прямоугольную форму. Кутки для концентрации улова представляют собой цилиндрические или конические мешки.
На Дальнем Востоке наибольшее распространение получили вентери кательного типа. Размеры крыльев и ловушек вентерей определяются условиями лова. В вентерях диаметр первого обруча принимается в пределах 1,5-3 м. Размер последующих обручей к кутку постепенно уменьшают, так чтобы они входили один в другой. Это делается для удобства транспортировки и работы с вентерями. Количество обручей в ловушке обычно не превышает 6-7, а расстояние между ними составляет от 0,5 до 0,75 диаметра обруча. В ряде случаев вход в ловушку делают прямоугольным, заменяя первый обруч рамой. Длина усынков должна равняться расстоянию между обручами или несколько превышать его. Широкой кромкой усынок прикрепляется к переднему обручу. Периметр входного отверстия в посадке принимается в пределах 0,2-0,7 длины окружности обруча. Оно расправляется специальными нитями-симками, которые завязываются за последующий обруч. В зависимости от количества симок отверстию усынки можно придать любую форму. В вентере без рамы высота крыла в приводной части и открылков превышает диаметр первого обруча. Поэтому дель открылков садят напервый обруч, а крыло пришивают к широкому основанию сетного клина, вставленного посредине входного усынка. Верхние подборы крыла и открылки крепятся на льду при помощи поводцов и деревянных планок. Нижние подборы оснащаются загрузкой из камней или другого материала. Между открылками поверху от первого обруча ставят козырек длиной 2-3 м. Боковые кромки козырька пришивают к верхним подборам открылков. Переднюю кромку козырька вмораживают в нижнюю кромку льда. В вентерях с рамой отпадает необходимость в применении сетных клиньев. Увеличение высоты рамы не требует обязательного изменения других частей вентеря.
Вентери устанавливают по одному или группами. Существует множество схем установки вентерей, которые определяются особенностями района промысла и распределением рыбы. На льду производят разметку вентеря в натуральную величину. Пробивают затяжную и рабочие майны, прогонные лунки. Протянув по всем направлениям прогонные веревки, подвозят и складывают на морской стороне рабочей майны вентерь кутком вниз, а раму, открылки и крыло сверху, причем открылки и крыло соединяются с рамой. Затем за концы прогонных веревок прикрепляют береговые кромки крыла и открылков и растягивают их подо льдом. При этом двор принимает рабочую форму. Затем кромки верхней подборы крыла и открылков закрепляют на льду. Прикрепив раму, открылки и крыло к конечным прорубям, вновь перебирают вентерь, но уже к береговой стороне рабочей майны, кутком кверху. Затем прогонную веревку соединяют с затяжкой и бочку вентеря затягивают и устанавливают на место.
Эксплуатация вентеря в процессе промысла складывается из выдержки его в воде, переборки и выливки улова, очистки майн ото льда и снега. Время выдержки вентеря в воде между переборками определяется опытным путем в зависимости от промысловой обстановки [2].
Качественные показатели рыбы сырца существенно могут ухудшаться за счет механических повреждений при освобождении орудий лова от улова. Снизить количество поврежденной рыбы возможно, нормируя количество рыбы в трале. При выливке улова из мешка трала применяют для этой цели не подъем стрелой, а вымывают рыбу забортной водой [5].
При использовании объячеивающих орудий лова (например, дрифтерных сетей) количество поврежденной рыбы снижают правильной настройкой сететрясных машин, а в некоторых случаях отказом от их использования.
На качество рыбы сырца наиболее существенное влияние оказывают температура ее хранения и высота слоя рыбы в тарных емкостях. Повышенная температура хранения и большая высота слоя рыбы ускоряет протекание автолитических процессов в сырье и вызывает его порчу [7].
С целью сохранения качества сырья его рекомендуют охлаждать и направлять на реализацию или переработку. При отсутствии такой возможности допускается непродолжительное хранение его без охлаждения.
Выловленную рыбу при температуре воздуха не более 10 0С допускается транспортировать в течение 2 ч. При этом ее предохраняют от воздействия прямых солнечных лучей, укрывая брезентом. Высота слоя мелкой рыбы и сельди не должна превышать 0,4 м. Для более крупных рыб высота слоя не должна превышать 0,7–0,8 м и зависит от вида рыбы и условий ее перевозки. При необходимости транспортирования и хранения более 2 ч ее необходимо охлаждать.
При хранении и транспортировании охлажденной рыбы необходимо регулярно контролировать температуру в трюме и состояние рыбы, откачивая воду из-под паел, не допуская ее накапливания и попадания на рыбу.
Сроки хранения и транспортирования охлажденной рыбы в первом и четвертом кварталах — до 12 сут, второй квартал — до 10 сут., третий квартал — до 7—8 сут. Мелкую рыбу хранят во льду на трое суток меньше, чем крупную. Предельные сроки хранения отдельных видов рыб устанавливают в зависимости от особенностей сырья, условий заготовки, транспортирования и хранения.
Использование биомицинового и хлорного льда позволяет значительно увеличить сроки хранения охлажденной рыбы [5].
3. Технохимическая характеристика трески
Треска (Gadus morhua macrocephalus). Обитатель морских вод умеренных и высоких широт северного полушария, тихоокеанская треска, хищная, типично морская стадная рыба, ведет миграционный образ жизни, но далеких перемещений не совершает.
Тихоокеанская треска достигает половозрелости на пятом году жизни, но вступают в нерест далеко не все половозрелые рыбы; нерест растянут, и происходит в разных районах в период с января по май.
Размеры и вес тихоокеанской трески в уловах изменяются в очень широких пределах (длина 24-115 см, вес 0,5-16 кг), но преобладают особи длиной 50—80 см и весом 3,5—5 кг. Это непостоянство размера и веса зависит от возрастного и полового состава косяков облавливаемой рыбы, различных темпов роста трески в отдельных районах, способа лова рыбы и т.п.
Между возрастом, длиной и весом тела у тихоокеанской трески имеется прямая зависимость, но в отдельных районах обитания темпы роста рыбы неодинаковы, поэтому корреляция веса и длины тела у тихоокеанской трески в различных районах ее лова выражается пучком кривых.
Как правило, в одном и том же районе лова средние размеры и вес трески, добываемой крючковой снастью, всегда выше, чем у трески, добываемой тралами.
У тихоокеанской трески самки всегда крупнее самцов по размерам и весу [3].
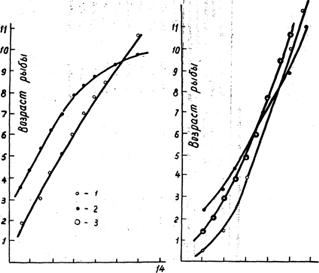
2 4 6 8 10 12 0,2. 0,4 0,6 0,8 1,0 1,2
Вес рыбы в кг Длина тела в м
Рис. 1. Зависимость размера и веса тихоокеанской трески от ее возраста. 1 — западное побережье. Камчатки, 2 — южные Курилы, 3 — юго-западное побережье Сахалина
Все это приводит к тому, что в одном и том же районе промысла средний вес и размер трески в уловах бессистемно колеблются как по месяцам, так и по отдельным годам.
Размеры и масса в уловах тихоокеанской трески изменяются в широких пределах и зависят от возраста и полового состава (длина 24-115 см, масса 0,5-16 кг). Преобладают особи длиной 50-80 см и массой 3-5 кг. Удельный вес неразделанной трески - 1,017-1,043. Насыпная масса в зависимости от размера - 900-1004 (кг/м3). Угол скольжения неразделанной рыбы для луженой жести 8-11°, алюминия 10-15°, оцинкованного железа 17-38°, кровельного железа 26-37°, гладкой резины 30-52°, строганого дерева -31-60 и бетона 33-68. Массовые соотношения частей тела трески представлены в табл. 1
Таблица 1
Массовые соотношения частей тела трески
| Части тела | Масса, кг | В % массы рыбы |
| Неразделанная рыба | 1,9 | 6,4 |
| Голова | 19,6-23,2 | |
|
В том числе: - жабры |
4,2-5,5 | |
|
Внутренности В том числе: - печень |
10,9-8,5 4,2-6,8 |
|
| Тушка | 51,3-57,9 | |
|
В том числе: - кожа - позвоночник - мышечная ткань без кожи |
2,0-4,0 5,0-6,4 44,2-47,6 |
|
|
Молоки Ястыки |
21,0-26,0 11,0-20,0 |
Весовые соотношения частей тела у тихоокеанской трески, добытой в разное время и в разных районах Дальнего Востока, изменяются незначительно.
Относительный вес половых желез у трески достигает максимума в январе-марте (молоки 21—26%-к весу самцов и ястыки 11—20% к весу самок); в июне (вскоре после нереста) вес половых желез не превышает 1,5-2% к весу рыбы, а в декабре вес молок достигает 11—17 и ястыков 20—23% к весу самцов и самок соответственно.
Таблица 2
Химический состав мяса тихоокеанской трески
| Район и период лова | Пределы содержания, % | |||
| вода | липиды | белок | Мин. вещества | |
| Зал. Петра Великого, август | 77,7-82,2 | 0,3-0,7 | 15,5-18,6 | 1,3-2,2 |
| Западное побережье Камчатки, июль-сентябрь | 79,5-82,0 | 0,3-0,6 | 15,8-17 | 1,0-1,8 |
| Охотское море, июль-сентябрь | 79,4-80,6 | 0,4-0,9 | 17,1-18,1 | 1,2-1,5 |
| Северное приморье, май- сентябрь | 80-81,9 | 0,3-0,5 | 16,8-17,3 | 0,8-1,3 |
|
Северный Сахалин, июль-август |
81-82,2 | 0,2-0,4 | 16,3-17,4 | 1,0-1,2 |
| Восточная часть Берингова моря, август-октябрь | 81,3-81,4 | 0,2-0,3 | 16,7-16,9 | 1,2-1,5 |
| Юго-восточная часть Берингова моря, июнь- август | 70,6-81 | 0,4-1,5 | 17,2-18,7 | 1,4-1,5 |
У тихоокеанский трески вес печени изменяется в зависимости отряда причин. С увеличением возраста рыбы увеличение массы печени происходит во много раз быстрее, чем увеличение веса рыбы (Кривобок). Так, например, при увеличении размера трески с 35 до 90 см вес рыбы возрастает в 14 раз (с 0,74 до 10,3 кг), а вес печени — в 34 раза (с 24 до 820 г). Поэтому с увеличением веса трески возрастает не только абсолютный, но и относительный вес печени.
Таблица 3
Химический состав частей тела тихоокеанской трески
| Части тела | Пределы содержания, % | |||
| влага | жир | белок | зола | |
| Голова | 78,2-82,4 | 0,7-0,9 | 14,3-15,4 | 7,4-8,3 |
| Плавники, хвост | 79,4-81,9 | 0,4-0,5 | 12,6-16,4 | 9,1-10,6 |
| Печень | 22,2-51,6 | 10,7-70,1 | 5,3-11,2 | 0,3-0,9 |
| Кишечник, желудок | 80,9-83,7 | 1,4-3,7 | 9,6-11,6 | 0,6-1,1 |
Относительная, масса печени находится в прямой зависимости от веса рыбы: у рыб весом 1,9—2,2 кг вес печени составляет 4,6—5,6%, а у рыб весом 4,6—5,6 кг — 5,6-6,5%.
У тихоокеанской трески имеется прямая связь между весом рыбы и относительным весом ее печени.
Имеющиеся у нас данные показывают, что существенных различий по выходу печени у тихоокеанской трески разных районов лова не имеется. По отдельным районам относительный вес печени изменяется в следующих пределах (в % к весу рыбы): северное Приморье — 3,2—6,8, западное побережье Камчатки — 5,1-—6,7; восточное побережье Камчатки 4—6, западное побережье южного Сахалина 5,4— 6,8, западное побережье северного Сахалина 5,8—7, восточное побережье южного Сахалина 5,8—11,6, юго-восточная часть Берингова моря 4,6—6,5 и Берингово море 3,2-6.
Можно отметить, что выход печени у самцов и самок примерно одинаков; но от весны к осени выход печени закономерно повышается.
У тихоокеанской трески относительная масса мяса изменяется от 38 до 48% к весу тела, без видимой зависимости от размеров и пола трески, а также периода и района ее лова.
Мясо тихоокеанской трески характеризуется малым содержанием жира и большим количеством влаги.
Можно отметить, что у половозрелой тихоокеанской трески после нереста (февраль—май) мясо становится более обводненным и менее вкусным. В результате интенсивного питания треска к осени достигает, нормальной упитанности.
У трески содержание влаги в мышцах у всех возрастных групп имеет сезонные колебания. По его данным, в апреле—мае треска имеет наиболее обводненное мясо (81% (влаги в мясе мелкой и 83,5% — в мясе крупной трески). Затем происходит постепенное снижение содержания влаги, которое в августе стабилизируется и держится на уровне 80-81,5% до января—февраля, когда влагосодержание в мясе начинает резко увеличиваться.
При сопоставлении известных в литературе данных с результатами анализов, выполненных, в ТИНРО, можно, установить, что химический состав мяса атлантической и тихоокеанской трески изменяется практически в одних и тех же пределах, однако верхние пределы содержания жира и белка в мясе тихоокеанской трески более высокие, чем в мясе атлантической.
Мясо трески отличается от мяса других видов семейства тресковых повышенным, содержанием небелкового азота (13—15% к общему азоту). Мясо трески содержит очень много ансерина (150 мг%), бетаина (102 мг%); окиси триметиламина {400-600 и до 1080 мг%), но в мясе нет гистидина и карнозина, a пo содержанию мочевины (1,8—2,7 мг%) и креатина (350—580 мг%) различий нет. В мясе трески несколько меньше свободных аминокислот, чем в мясе других видов донных рыб, причем в составе свободных аминокислот меньше незаменимых, особенно метионина, лизина и циклических аминокислот; среди заменимых низкое содержание моноаминокислот, аргинина, пролина и тирозина.
В мясе трески присутствуют все важные водорастворимые витамины (γ%): B1 50—95, В2 110—830, Bс 10—12, B12 0,7—1,4, РР 1000—3200 и пантотеновой кислоты 50—200.
В мясе трески обнаружен полноценный комплекс элементов, входящих в состав минеральных веществ мяса (мг % к весу сырого вещества): калий 220—240, Кальций 11—30, магний 24-140, фосфор 170—230, железо 0,5—1,1, цинк 6—10 (γ %), йод 90—120, марганец 50—60, медь 60—70, кобальт 20—30, молибден 6—10, бром 0,5—0,6. Ткани частей тела тихоокеанской трески содержат мало жира, за исключением печени.
В печени сосредоточивается практически весь жировой запас организма рыбы. Масса накапливающегося в печени жира составляет от 1,7 до 6,1% к весу ее тела.
Содержание жира в печени и витамина А в печеночном жире изменяется в больших пределах, причем это непостоянство проявляется во всех районах лова тихоокеанской трески.
Окраска печени связывается с содержанием в ней жира: печень с высоким содержанием жира имеет желтую или светло-кремовую окраску; с уменьшением содержания жира в окраске появляются красно-коричневые тона.
Установлено, что содержание жира и витамина А в печени трески зависит от целого ряда причин биологического характера. Влияние этих причин проявляется в сложных взаимосочетаниях, что в. ряде случаев затрудняет выявление закономерностей, которым подчиняются изменения содержания жира и витамина А в печени.
К причинам, отчётливо влияющим на содержание жира и витамина А в печени, следует отнести возраст и упитанность рыбы, а также сезон и район ее лова. На примере трески, добытой в июле в районе восточного побережья Камчатки .(Олюторский и Краноцкий заливы), можно было установить, что у рыб старше 4 лет содержание витамина А в печени и печеночном жире оказывается наибольшим.
Содержание жира в печени и витамина А в жире возрастает с увеличением веса трески. Так, у трески весом 1,5—2,5 кг в 1 г печеночного жира содержалось 2010—4610 и. е. витамина А, у рыб весом 2,5-4 кг печеночного жира было 4670-4960 и.е., а у трески 5-6 кг -7000-7500 и.е. Содержание жира в печени рыб весом до 2,5 кг изменялось oт l6 до 20%, а у рыб более 2,5 кг вecoм от 22 дo 32%.
Более значительные изменения концентрации витамина А в печеночном жире обнаружены у беринговоморской трески, добытой в период август-октябрь. Так, у рыб весом до 1,5 кг содержание жира в печени изменялось от 13,1 до 27,8%, а содержание витамина A в 1 г печеночного жира от 510 до 2480 и. е. У рыб весом 6—7,2 кг содержание жира от 10,7 до 30,2%, а витамина А от 2200 до 17510 и. е. на 1 г печеночного жира[3].
Таким образом, с увеличением в уловах количества трески и старших возрастов следует ожидать более высокого выхода жира и витамина A.
Для трески одинакового веса (возраста), пойманной в Олюторском заливе в июле—августе, было установлено, что по сравнению c самцами самки имеют более жирную печень(21,7-34.4 и 40,6—45,4% жира соответственно), но в печеночном жире последних витамина А несколько меньше-1490-3490 и 1480—1610 и.е. соответственно. Поэтому у самцов на 1 кг веса тела накапливается несколько больше витамина А, чем у самок (37740—67410 и 41040—52930 и.е. соответственно. Поэтому при увеличении в уловах количества крупных самцов можно ожидать некоторого уменьшения выхода печеночного жира и повышения в жире витамина А.
У тихоокеанской трески обнаруживаются сезонные колебания в содержании жира и витамина А в печени. У половозрелой трески содержание жира к периоду нереста заметно снижается и достигает минимума в посленерестовый период. В период нагула скорость восполнения запасов жира в печени находится в прямой зависимости от интенсивности питания.
Содержание жира в печени достигает максимума в сентябре, а наиболее, высокое содержание витамина А обнаруживается значительно раньше — в июле.
У трески из Олюторскогр залива содержание жира в печени с 21,7—34,4% в июле возросло до 38—51% в сентябре, а среднее содержание витамина А в печени и в печеночном жире увеличилось с 740 до 1380 и с 2510 до 3100 и. е. соответственно.
Процессы продуцирования и накопления жира, а также витамина А в печени связаны, очевидно, с интенсивностью нагула.
Кроме печени витамин А обнаружен в тканях кишечника, желудка и пилорических придатков. Относительный вес желудка вместе с кишечником изменяется от 4,8 до 12,6% к весу рыбы, а содержание жира от 1,2 до 4%. В 1 г тканевого жира содержится от 770 до 47230 и. е. витамина А. В пилорических придатках содержание жира изменяется от 1,3 до 2,9%, а в 1 г тканевого жира содержится от 1280 до 28000 и. e. витамина А.
Таким образом, в организме трески основная часть (70—75%) запаса витамина А депонируется в печени.
В тканях тресковой печени обнаружили витамин B2 (790—1540 γ %), пантотеновую кислоту (230—330 γ %), тиамин (65—105), Вс (80—85) и очень высокое содержание витамина В12 (100-480 γ %).
Извлеченный из безупречно свежей тресковой печени жир имеет соломенно-желтую окраску, приятные органолептические свойства и низкую кислотность (0,1-0,3).При хранении печени до вытопки жира за счет деятельности тканевых липаз увеличивается кислотность жира в прямой зависимости от продолжительности и температуры хранения, причем гидролиз жира идет быстрее в случае, если печень находится в брюшной полости рыбы. Taк, при температуре 13—20°С двухсуточное хранение неразделанной трески ведет к увеличению кислотности печеночного жира до 5,9, в этих же условиях печень, извлеченная перед хранением из брюшной полости, имеет жир с кислотностью не выше 3,5-4 мг КОН 1 г жира. Повышение температуры при, хранении ведет к ускорению гидролиза жира. Так, при одинаковой. :продолжительности хранения кислотность печеночного жира при 18-24°С в1,5—2 раза выше, чем при температуре 0-1°С.
Задержка печени перед обработкой сопровождается усилением окраски жира и специфического запаха и вкуса. Это связывается с накоплением в жире не только продуктов его гидролиза и окисления, но и азотистых оснований.
В целях сохранения природных свойств печеночного жира печень должна извлекаться из брюшной полости возможно быстрее после подъема трески из воды и без задержек направляться для вытопки жира. Вынужденное хранение рыбы или печени должно проходить только при охлаждении.
Физико-химические свойства печеночных жиров тихоокеанской трески изменяются в небольших пределах.
Тресковый печеночный жир имеет число Рейхерта-Мейселя 0,4—1, число Генера 93—96,5.
Неомыляемые вещества в основном представлены холестерином, в меньшей мере лецитином и липохромами. В жирах промышленной выработки присутствуют азотистые основания, в частности триметиламин. Чем интенсивнее окраска жира и чем сильнее в жире проявляются неприятный запах и вкус, тем больше в нем содержится азотистых оснований. Светло окрашенные жиры имеют кислотность не выше 0,5, темно-жёлтые от 0,5 до 2,5, желто-коричневые 2—5 и коричневые до 30-38 (мг КОН 1 г жира).
Таблица 3
Аминокислотный состав белков мышечной ткани трески
| Аминокислоты | Содержание, % белка |
|
|||
| незаменимые | заменимые |
|
|||
| 1 | 2 | 3 |
|
||
| Монокарбоновые | Валин | 5,3 | Глицин | 1,0 |
|
| Лейцин | 8,0 | ||||
| Изолейцин | 5,5 | Алании | - | ||
| Треонин | 5,0 | Серии | - | ||
| Серосодержащие | Метионин | 3,0 | |||
| Диаминокислоты | Лизин | 9,0 | |||
| Циклические аминокислоты | Фенилаланин | 3,8 | Аргинин | 6,0 | |
| Гистидин | 2,0 | Пролин | - | ||
| Триптофан | 1,0 | ||||
В печеночном тресковом жире содержится 8,6-14%. твердых, жирных кислот (йодное число 13-17 и титр 51,2—52,2° С) и 85—87% жидких ненасыщенных (йодное_число 166,1-171,6, η -1,4729). Твердые жирные кислоты в основном представлены пальмитиновой (6,5—13%) и миристиновой (3,5—6,5%) кислотами и незначительным (до 0,5%) количеством стеариновой кислоты.
В составе ненасыщенных жирных кислот преобладают кислоты C18 (25—31%) с ненасыщенностью — 2,8—2,9 и С20 (26—31,5%) с ненасыщеннсстью - 6—6,1. Сильноненаеыщённых кислот (6,4-6,9) С22 содержится 10—14%. Количество ненасыщенных (2,0—2,9) кислот C16 не превышает 15—20%, а кислот С14 — не более 0,5%.
Значительное содержание в жире сильноненасыщенных, жирных кислот делает тресковый печеночный жир неустойчивым к действию кислорода воздуха.
При охлаждении трескового печеночного жира выделяется кристаллическая фракция, которая отделяется от жидкой части путем фильтрации и прессования; выход твердого “стеарина” составляет 4—8% к весу жира. Твердая фракция (температура плавления 37—38е) содержит 43—44% твердых жирных кислот (йодное числ 10—11, титр 52,8—53,2°С), 49% непредельных (йодное число 105, η —1,4636) и l,5-2,5% неомыляемых веществ.
Исследования процесса вытопки жира из печени тихоокеанской трески показали, что выход жира колеблется от 25,2 до 38% (среднее 30%) к весу печени, причем содержание витамина А в жире колеблется от 1090 до 4240 (среднее 2780) и. е. в 1г.
Установлено, что наибольший выход жира получается при первом сливе, причем по содержанию витамина А жир первого и второго сливов и жир отпрессованный существенных различий не имеет, но отличается по окраске и кислотности [3].
Производство, транспортирование и хранение охлажденной трески проводится на основе ГОСТ 814- 96. Охлажденная рыба должна быть изготовлена в соответствии с требованиями настоящего стандарта по технологической инструкции, с соблюдением санитарных норм и правил, утвержденных в установленном порядке.
По видам разделки охлажденную треску подразделяют на:
1.Неразделанную - рыба в целом виде.
2. Потрошеная с головой - рыба, разделанная по брюшку между грудными плавниками от калтычка до анального отверстия или на 1,5- 2,0 см далее; калтычек может быть перерезан; голова, внутренности удалены, сгустки крови зачищены; вырезана зона анального отверстия с частичным подрезом анального плавника, оставлены недоразвитые икра или молоки.
В охлажденной рыбе не должно быть живых гельминтов и их личинок, опасных для здоровья человека.
Сырье и материалы (рыба живая, рыба- сырец, лед), используемые для изготовления охлажденной рыбы должны быть не ниже первого сорта.
Охлажденную рыбу упаковывают в тару со льдом. Массовая доля льда в момент выпуска с предприятий должна быть не менее 50% по отношению к массе рыбы [4].
Охлажденную рыбу упаковывают:
- в ящики деревянные по ГОСТ 13356 предельной массой продукта 75 кг;
- в бочки сухотарные по ГОСТ 8777 вместимостью не более 150 дм3.
Тара для упаковывания охлажденной рыбы должна быть плотной, чистой, без постороннего запаха.
Деревянные ящики между дощечками дна должны иметь просветы шириной не более 0,5 см, а в днищах бочек- отверстия для стока воды, образующейся от таяния льда.
Рыбу длиной менее 30 см укладывают в тару насыпью с разравниванием по слоям.
Рыбу длиной более 30 см укладывают в тару ровными рядами спинкой вверх.
На дно тары и на каждый ряд (слой) рыбы насыпают слой чистого мелкодробленого льда.
В каждой упаковочной единице должна быть рыба одного наименования, вида разделки, одной размерной группы.
Деревянные ящики должны быть плотно забиты, а для иногородних перевозок, кроме того, по торцам скреплены стальной упаковочной лентой по ГОСТ 3560 или стальной проволокой по ГОСТ 3282.
Бочки с рыбой должны быть плотно укупорены.
Маркируют тару с продукцией по ГОСТ 7630.
Правила приемки осуществляют по ГОСТ 7631. Контроль за содержанием токсичных элементов, пестицидов и гистамина осуществляют в соответствии с порядком, установленным производителем продукции по согласованию с органами государственного санитарно- эпидемиологического надзора [4].
Таблица 5
Схема направления трески на производство пищевых продуктов [2]
| Мороженые | Копченые | Консервы | ||||||||||
| Вид | Живые или охлажденные | неразделанные потрошеная | филе | фарши | Соленые и пресервы | Сушеные | Вяленые | горячее | холодное | Кулинария | натуральные | закусочные |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 |
| Тихоокеанская треска | + | – | +++ | + | ++ | – | – | +++ | ++ | +++ | – | +++ |
(-)- не используется
(+)- допускается производить продукцию
(+++)- основное направление в использовании
траловый треска химический пищевой
Таблица 6
Схема направления отходов от разделки трески на производство пищевых, кормовых, технических продуктов и биологически активных веществ [2]
| Продукты | |||||
| Вид | Части тела | пищевые | кормовые | технические | биологически активные вещества |
| 1 | 2 | 3 | 4 | 5 | 6 |
| Тихоокеанская треска |
Головы Печень Плавательный пузырь Прочие отходы |
Стерилизованные консервы Стерилизованные консервы- |
Кормовая мука – Кормовая мука |
- Жир Рыбный клей |
|
Заключение
Тихоокеанская треска имеет вальковатое тело, широкую голову и длинный усик на нижней челюсти. Голова у нее относительно крупнее и шире, чем у атлантической трески. Она сильно отличается от атлантической трески строением рожкообразных выростов переднего конца плавательного пузыря, которые у нее гораздо короче, чем у атлантической.
В водах Приморья треска встречается повсеместно. Широко представлена в Японском, Охотском и Беринговом морях, а также у берегов Северной Америки на юг до Орегона, в тихоокеанских водах Японии, в Желтом море, у берегов Кореи и Китая.
Лов трески возможен круглый год, однако в Северном Приморье между м. Олимпиады и б. Нельма, для удобного и ярусного промысла наиболее результативны июль-август, когда треска откармлипается вблизи берегов на глубинах 30-75 м. Запасы трески в водах Приморья недоиспользуются. В последнее десятилетие её вылов здесь колебался в пределах 0,35-2,20 тыс. т. в год. Добывается крючковыми снастями (ярусами и удочками) и тралящими орудиями лова (снюрреводами и тралами). При подходах к берегу попадается в ставные невода.
Мясо трески отличается от мяса других видов семейства тресковых повышенным, содержанием небелкового азота (13—15% к общему азоту). В мясе трески несколько меньше свободных аминокислот, чем в мясе других видов донных рыб, причем в составе свободных аминокислот меньше незаменимых, особенно метионина, лизина и циклических аминокислот; среди заменимых низкое содержание моноаминокислот, аргинина, пролина и тирозина в мясе трески обнаружен полноценный комплекс элементов, входящих в состав минеральных веществ мяса (мг% к весу сырого вещества): калий 220—240, Кальций 11—30, магний 24-140, фосфор 170—230, железо 0,5—1,1, цинк 6—10 (γ %), йод 90—120, марганец 50—60, медь 60—70, кобальт 20—30, молибден 6—10, бром 0,5—0,6. Ткани частей тела тихоокеанской трески содержат мало жира, за исключением печени.
Треска пользуется широким спросом на рынках всех европейских стран. Она доставляется в парном или замороженном виде, особенно ценится приготовленное из нее филе. Жир из тресковой печени богат витаминами А и D и представляет собой ценный медицинский препарат, особенно для детей, против заболевания рахитом. В Норвегии заготавливают также икру трески, которая продается в небольших тюбиках и заслуженно пользуется большим спросом во многих странах Европы.
Список литературы
1. Новиков В.М. Рыбы Приморья.- Владивосток: Дальрыбвтуз, 2003.- 254с.
2. Новиков В.М. Справочник по прибрежному рыболовству.- Владивосток: Дальрыбвтуз, 1999.- 262с.
3. Кизеветтер И.Н. Технохимическая характеристика рыб Дальневосточного бассейна.- Владивосток: ДВОРАН, 1973.- 424с.
4. Касьянов Г.И. Технология переработки рыбы и морепродуктов.- Ростов-на-Дону: изд. центр «МарТ»,2001.- 273с.
5. Андрусенко П.И. Технология рыбных продуктов.- М.: Агропромиздат, 1989.- 315с.
6. Дацун В.М. Вторичные ресурсы рыбной промышленности.- М.: Колос, 1985.- 96с.
7. Перебейнос А.В. Новые кормовые продукты из отходов переработки морских гидробионтов.- Владивосток: ДВГУ, 1995.- 140с.
8. Сафронова Т.М. Органолептическая оценка рыбной продукции.- М.: Агропромиздат,1985.- 191с.